- Trang chủ
- Sách y học
- Sinh lý y học
- Sự giải phóng năng lượng từ Glucose cho cơ thể theo con đường đường phân
Sự giải phóng năng lượng từ Glucose cho cơ thể theo con đường đường phân
Cách quan trọng nhất để giải phóng năng lượng từ glucose là khởi động con đường đường phân, sản phẩm cuối cùng sau đó được oxy hóa để cung cấp năng lượng.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Vì sự oxy hóa hoàn toàn 1 mol glucose giải phóng ra 686,000 calo và chỉ cần 12,000 calo để tổng hợp nên 1 mol ATP, năng lượng có thể bị lãng phí nếu glucose bị phân hủy toàn bộ thành nước và CO2 trong khi chỉ tạo ra một phân tử ATP. May mắn là, tế bào trong cơ thể chứa những enzym đặc biệt làm cho phân tử glucose chia cắt từng phần nhỏ trong một thời điểm và qua nhiều bước liên tiếp, vì thế năng lượng được giải phóng ra thành từng phần nhỏ để tạo nên một phân tử ATP tại một thời điểm, do đó mỗi mol glucose được tế bào chuyển hóa tạo ra tổng cộng 38 mol ATP.
Đường phân (Glycolysis) - Glucose thành Pyruvic Acid
Cho đến nay cách quan trọng nhất để giải phóng năng lượng từ glucose là khởi động con đường đường phân. Sản phẩm cuối cùng của quá trình đường phân sau đó được oxy hóa để cung cấp năng lượng. Đường phân nghĩa là chia cắt một phân tử glucose để tạo thành hai phân tử pyruvic acid.
Đường phân xảy ra bởi 10 phản ứng hóa học liên tiếp. Mỗi bước được xúc tác bởi ít nhất một enzym đặc hiệu. Chú ý rằng ban đầu, glucose được chuyển thành fructose-1,6-diphosphate và sau đó chia thành hai phân tử có 3-carbon , glyceraldehyde-3-phosphate, chúng sẽ được chuyển thành pyruvic acid thông qua 5 bước nữa.
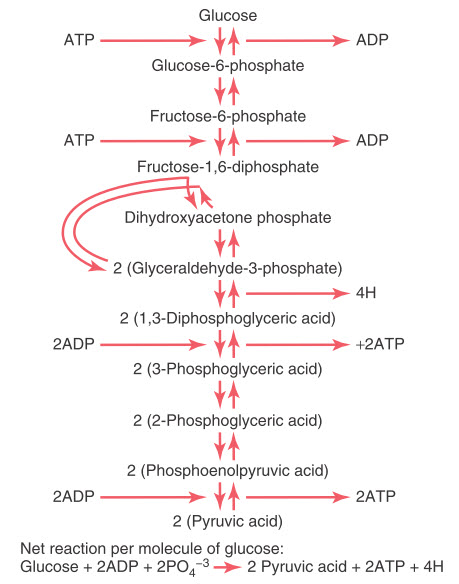
Hình. Chuỗi các phản ứng hóa học gây ra quá trình đường phân.
Sự tạo thành ATP trong quá trình đường phân. Mặc dù có nhiều phản ứng hóa học trong quá trình đường phân, chỉ có một phần nhỏ năng lượng tự do trong phân tử glucose được giải phóng tại hầu hết các bước. Mặc dù vậy, trong giai đoạn giữa 1,3-diphosphoglyceric acid và 3-phosphoglyceric acid, và giai đoạn giữa phosphoenolpyruvic acid và pyruvic acid, một phần của năng lượng được giải phóng lớn hơn 12,000 calo mỗi mol, số lượng cần thiết để tổng hợp nên ATP, và phản ứng tổng hợp ATP được hình thành. Do đó, tổng cộng 4 mol ATP đã được tạo ra từ 1 mol fructose-1,6-diphosphate được phân chia thành pyruvic acid.
Tuy nhiên, cần 2 mol ATP để phosphoryl hóa glucose ban đầu thành fructose-1,6-diphosphate trước khi quá trình đường phân bắt đầu. Do đó, số lượng thực tế thu được của cả quá trình đường phân chỉ là 2 mol ATP cho mỗi mol glucose được sử dụng. Số lượng này tương đương 24000 calo đã được chuyển thành ATP, nhưng trong quá trình đường phân, tổng cộng có 56,000 calo đã mất từ phân tử glucose ban đầu, thành ra hiệu suất của toàn bộ quá trình tổng hợp ATP chỉ là 43%. 57% năng lượng còn lại mất đi dưới dạng nhiệt.
Chuyển hóa Pyruvic Acid thành Acetyl Coenzyme A
Giai đoạn tiếp theo trong sự thoái hóa của glucose là chuyển hóa qua hai bước của hai phân tử pyruvic acid thành hai phân tử acetyl coenzyme A (acetyl-CoA) bằng các phản ứng sau:
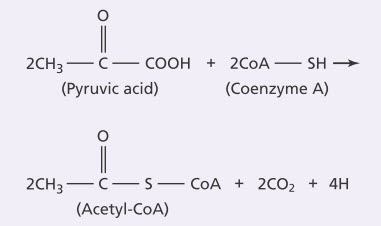
Hai phân tử CO2 và 4 nguyên tử hydro được giải phóng từ phản ứng này, trong khi phần còn lại của hai phân tử pyruvic acid liên kết với coenzyme A, là một dẫn xuất của vitamin B5 (pantothenic acid), để tạo thành hai phân tử acetyl-CoA. Trong chuyển hóa này, không có ATP nào được tạo ra, nhưng tới 6 phân tử ATP sẽ được tạo thành khi 4 nguyên tử hydro vừa giải phóng được oxy hóa sau đó và sẽ được đề cập sau đây.
Chu trình Citric Acid (Chu trình Krebs)
Giai đoạn tiếp theo của quá trình thoái hóa glucose gọi là chu trình citric acid (còn được biết dưới cái tên chu trình tricarboxylic acid hoặc chu trình Krebs nhằm vinh danh Hans Krebs - người đã khám phá ra chu trình này). Chu trình citric acid là một chuỗi các phản ứng hóa học trong đó phần acetyl của phân tử acetyl-CoA được thoái hóa thành CO2 và nguyên tử Hydro. Tất cả những phản ứng này đều xảy ra ở trong chất nền của ty thể. Sự giải phóng nguyên tử hydro làm tăng thêm số lượng nguyên tử sẽ bị oxy hóa, giải phóng ra một năng lượng cực lớn để hình thành nên ATP.
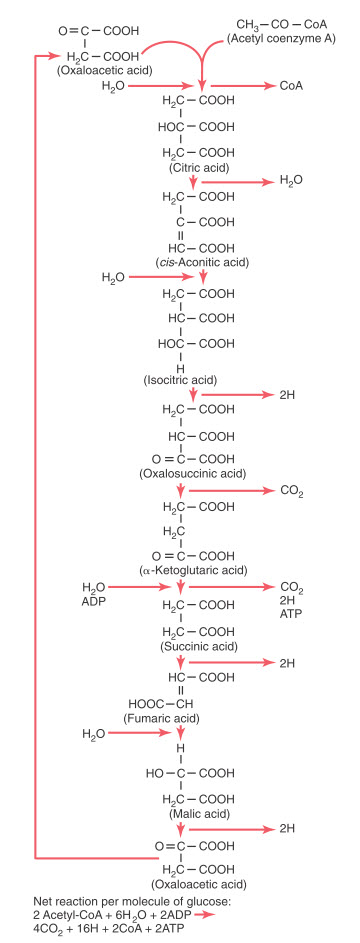
Hình. Phản ứng hóa học của chu trình axit xitric, cho thấy sự giải phóng khí cacbonic và một số nguyên tử hydro trong chu trình.
Các giai đoạn khác nhau của các phản ứng hóa học trong chu trình citric acid. Ở bên trái là các chất tham gia thêm vào phản ứng, còn sản phẩm của phản ứng được thể hiện ở bên phải. Chú ý rằng vị trí cao nhất trong sơ đồ là nơi chu trình bắt đầu với oxaloacetic acid, và khi chuỗi phản ứng kết thúc, oxaloacetic acid được tái tổng hợp một lần nữa. Do đó chu trình có thể được lặp lại nhiều lần.
Trong giai đoạn khởi đầu của chu trình citric acid, acetyl-CoA gắn với oxaloacetic acid tạo thành citric acid. Phần coenzyme A của acetyl-CoA được giải phóng và tái sử dụng nhiều lần để tổng hợp thêm một số lượng lớn acetyl-CoA từ pyruvic acid. Còn lại, toàn bộ phần acetyl được sử dụng để tạo nên phân tử citric acid. Trong các giai đoạn kế tiếp của chu trình citric acid, có rất nhiều phân tử nước tham gia vào quá trình, trong khi carbon dioxide và nguyên tử hydro được giải phóng ra ở các giai đoạn khác.
Kết quả cuối cùng của toàn bộ chu trình citric acid được giải thích, chứng minh rằng với mỗi phân tử glucose ban đầu sau khi được chuyển hóa ra được hai phân tử acetyl-CoA sẽ đi vào chu trình citric acid cùng với 6 phân tử nước. Những phân tử này sau đó sẽ thoái hóa thành 4 phân tử carbon dioxide, 16 nguyên tử hydro và 2 phân tử coenzyme A. Hai phân tử ATP được tạo thành theo cách thức mô tả dưới đây.
Sự hình thành ATP trong chu trình Citric Acid. Chu trình citric acid tự bản thân nó không giải phóng ra được nhiều năng lượng; chỉ một phân tử ATP được tạo thành trong duy nhất một phản ứng hóa học - trong quá trình từ α-ketoglutaric acid thành succinic acid. Do đó với mỗi phân tử glucose được chuyển hóa sẽ ra hai phân tử acetyl-CoA tham gia vào chu trình citric acid, mỗi phân tử chỉ tạo ra một phân tử ATP thành ra có tổng cộng là 2 phân tử ATP đã được hình thành.
Chức năng của enzym Dehydrogenases và coenzym Nicotinamide Adenine Dinucleotide trong việc giải phóng nguyên tử Hydro ở chu trình Citric Acid. Như đã nhấn mạnh tại một số điểm trong phần thảo luận này, nguyên tử hydro được giải phóng từ các phản ứng khác nhau trong chu trình citric acid - 4 nguyên tử hydro từ quá trình đường phân, 4 trong quá trình tổng hợp acetyl-CoA từ pyruvic acid, và 16 trong chu trình citric acid; do đó tổng cộng là 24 nguyên tử hydro được giải phóng từ phân tử glucose ban đầu. Mặc dù vậy, nguyên tử Hydro không tồn tại tự do trong dịch nội bào. Thay vào đó chúng được giải phóng ra từng đôi, và trong trường hợp này là nhờ xúc tác của một enzym đặc hiệu có tên là dehydrogenase. 20 trong tổng số 24 nguyên tử hydro ngay lập tức liên kết với nicotinamide adenine dinucleotide (NAD+), một dẫn xuất của vitamin B3 (niacin), theo phản ứng:
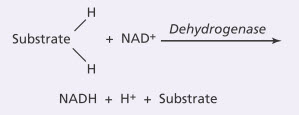
Phản ứng này sẽ không xảy ra nếu không có enzym dehydrogenase hoặc không có NAD+ làm nhiệm vụ như một chất mang hydro. Cả ion H+ tự do và hydro gắn với NAD+ sau đó đều tham gia vào các phản ứng oxy hóa để tổng hợp ra một số lượng lớn ATP, như đề cập sau đây.
4 nguyên tử hydro còn lại giải phóng từ quá trình thoái hóa của glucose - chính xác là trong chu trình citric acid ở giai đoạn chuyển từ succinic thành fumaric acid - liên kết với một enzym dehydrogenase đặc hiệu nhưng sau đó không được giải phóng vào NAD+. Thay vào đó chúng đi trực tiếp từ enzym dehydrogenase vào quá trình oxy hóa.
Chức năng của enzym Decarboxylases trong việc giải phóng carbon dioxide. Nhắc lại những phản ứng hóa học trong chu trình citric acid, cũng như quá trình hình thành acetyl-CoA từ pyruvic acid, chúng ta thấy rằng có 3 giai đoạn giải phóng ra carbon dioxide. Để giải phóng carbon dioxide, có những enzyme đặc hiệu gọi là decarboxylases, làm nhiệm vụ cắt carbon dioxide ra từ cơ chất. carbon dioxide sau đó được hoàn tan vào trong dịch của cơ thể và vận chuyển đến phổi, tại đó nó được đào thải ra khỏi cơ thể.
Bài viết cùng chuyên mục
Tăng huyết áp có angiotensin tham gia: gây ra bởi khối u tiết renin hoặc thiếu máu thận cục bộ
Một khối u của các tế bào cận cầu thận tiết renin xuất hiện và tiết số lượng lớn của renin, tiếp theo, một lượng tương ứng angiotensin II được hình thành.
Sợi thần kinh: sự tương quan về trạng thái
Một số tế bào thần kinh trong hệ thần kinh trung ương hoạt động liên tục vì ngay cả trạng thái kích thích bình thường cũng trên ngưỡng cho phép.
Bôi trơn bảo vệ và tầm quan trọng của chất nhày trong đường tiêu hóa
Chất nhày có khả năng khiến cho sự trượt của thức ăn trong đường tiêu hóa rất dễ dàng và ngăn cản sự trầy xước cơ học hoặc sự phân hủy hóa học cho lớp biểu mô.
Tiểu thể Pacinian: điện thế nhận cảm và ví dụ về chức năng của receptor
Tiểu thể Pacinian có một sợi thần kinh trung tâm kéo dài suốt lõi tiểu thể. Bao quanh sợi thần kinh trung tâm này là các lớp vỏ bọc khác nhau xếp đồng tâm, và do vậy, sự đè ép ở bất kì vị trí nào bên ngoài tiểu thể sẽ kéo giãn.
Giải phẫu chức năng của khu liên hợp vỏ não
Những khu vực liên hợp cũng có phân hóa chuyên môn riêng của nó. Các khu vực liên hợp quan trọng bao gồm: (1) khu liên hợp đỉnh- chẩm, (2) khu liên hợp trước trán, và (3) khu liên hợp hệ viền.
Hình thành acid acetoacetic trong gan và sự vận chuyển trong máu
Các acid acetoacetic, acid β-hydroxybutyric, và acetone khuếch tán tự do qua màng tế bào gan và được vận chuyển trong máu tới các mô ngoại vi, ở đây, chúng lại được khuếch tán vào trong tế bào.
Sự điều hòa nồng độ canxi cơ thể người
Ngay khi cơ chế canxi dễ trao đổi trong xương kiểm soát nồng độ canxi dịch ngoại bào, cả hệ PTH và calcitonin đều phản ứng. Chỉ trong 3-5 phút sau sự tăng cấp tính của ion canxi, tốc độ tiết PTH giảm.
Phức bộ QRS: hình dạng giãn rộng bất thường
Phức bộ QRS được xem là không bình thường khi kéo dài ít hơn 0,09s; khi nó giãn rộng trên 0,12s- tình trạng này chắc chắn gây ra bởi bệnh lý block ở 1 phần nào đó trong hệ thống dẫn truyền của tim.
Duy trì huyết áp động mạch bình thường: vai trò của hệ thống Renin-Angiotensin mặc dù có biến đổi lớn lượng muối vào
Hệ thống renin-angiotensin có lẽ là hệ thống mạnh mẽ nhất của cơ thể, làm thay đổi nhỏ huyết áp động mạch khi có lượng muối nhập vào dao động lớn.
Kiểm soát hoạt động của trung tâm hô hấp và các tín hiệu ức chế hít vào
Tính tới thời điểm này, đã biết về các cơ chế cơ bản tạo ra hiện tượng hít vào và thở ra, nhưng cũng rất cần tìm hiểu xem làm thế nào cường độ tín hiệu điều hòa có thể làm tăng hoặc giảm thông khí theo như cầu của cơ thể.
Sự trưởng thành và thụ tinh của trứng: quyết đinh giới tính thai nhi
Sau khi phóng tinh dịch vào trong âm đạo trong quá trình giao hợp, một số tinh trùng sẽ di chuyển trong vòng 5 đến 10 phút theo hướng đi lên từ âm đạo đến tử và vòi tử cung, đến đoạn bóng của vòi tử cung.
Nguyên nhân gây ngoại tâm thu: rối loạn nhịp tim
Ngoại tâm thu thường xuyên gặp trong thông buồng tim, ngoại tâm thu cũng xảy ra khi đứa catheter vào trong buồng thất phải và chén ép nội tâm mạc.
Sự tổng hợp Carbohydrates từ Proteins và chất béo - Quá trình tân tạo glucose
Khi kho dự trữ carbohydrates của cơ thể giảm xuống dưới mức bình thường, một lượng vừa phải glucose có thể được tổng hợp từ amino acids và phần glycerol của chất béo.
Cơ sở giải phẫu của hấp thu ở ruột
Bề mặt hấp thu của niêm mạc ruột non, thể hiện nhiều nếp gấp được gọi là nếp vòng, chúng tăng diện tích hấp thu của niêm mạc khoảng 3 lần, chúng thường nhô lên 8mm vào lòng ruột.
Điện thế tai trong: phản ánh nồng độ cao kali và nồng độ thấp natri
Điện thế được duy trì liên tục giữa nội dịch và ngoại dịch, với bản dương bên trong và bản âm bên ngoài thang giữa. Nó được gọi là điện thế tai trong.
Tăng huyết áp: gây ra bởi sự kết hợp của tăng tải khối lượng và co mạch
Tăng huyết áp di truyền tự phát đã được quan sát thấy ở một số chủng động vật, bao gồm các chủng chuột, thỏ và ít nhất một chủng chó.
Sinh lý điều trị đái tháo đường
Insulin có một số dạng. Insulin "Thường xuyên" có thời gian tác dụng kéo dài 3-8 giờ, trong khi các hình thức khác của insulin được hấp thụ chậm từ chỗ tiêm và do đó có tác dụng kéo dài đến 10 đến 48 giờ.
Calcitonin và canxi huyết
Sự kích thích chính tiết calcitonin là việc tăng nồng độ ion canxi dịch ngoai bào. Ngược lại, sự tiết PTH được kích thích bởi nồng độ canxi giảm.
Sự nhạy cảm quá mức của các cơ quan chịu sự chi phối của hệ giao cảm và phó giao cảm sau khi loại bỏ dây thần kinh
Nguyên nhân của sự nhạy cảm quá mức mới chỉ được biết một phần. Là số lượng các receptor trên màng sau synap của các tế bào đích đôi khi tăng lên vài lần ở vị trí noradrenalin hoặc acetylcholin không còn được giảI phóng vào các synap.
Điều hòa gen trong cơ thể người
Ở động vật có nhiều loại tế bào, mô, cơ quan khác nhau, các điều hòa biểu hiện gen khác nhau cũng cho phép nhiều loại tế bào khác nhau trong cơ thể thực hiện các chức năng chuyên biệt của chúng.
Hô hấp trong tập luyện thể thao
Có một mối quan hệ tuyến tính. Cả tiêu thụ oxy và tổng thông khí phổi tăng gấp khoảng 20 lần từ trạng thái nghỉ ngơi và cường độ tập luyện tối đa ở các vận động viên được tập luyện tốt.
Đường kính đồng tử: cơ chế quang học của mắt
Khả năng hội tụ tốt nhất có thể đạt được là khi lỗ đồng tử co nhỏ tối đa. Lí giải cho điều đó, với một lỗ đồng tử rất nhỏ, gần như tất cả các tia sáng đi qua trung tâm của hệ thấu kính của mắt.
Thị giác từ hai mắt: hợp nhất các hình ảnh
Sự tương tác diễn ra giữa các neuron ở vỏ não để kích thích can thiệp vào các neuron đặc hiệu khi hai hình ảnh thị giác không được “ghi nhận” - nghĩa là, không “hợp nhất” rõ ràng.
Chức năng sinh lý của oxytocin
Oxytocin làm co tử cung khi mang thai, khiến vú bài xuất sữa, Hormone oxytocin, kích thích mạnh mẽ co tử cung khi mang thai, nhất là trong giai đoạn cuối của thai kì.
Chức năng của testosterone
Testosterone được tiết ra đầu tiên ở các tế bào mầm rãnh sinh dục và sau đó là tinh hoàn của thai nhi chịu trách nhiệm trong sự phát triển các đặc điểm cơ thể nam giới, bao gồm cả sự hình thành dương vật và bìu chứ không phải là âm vật và âm đạo.
