- Trang chủ
- Sách y học
- Sinh lý y học
- Cơ chế co cơ trơn
Cơ chế co cơ trơn
Một đặc tính quan trọng khác của cơ trơn, đặc biệt là loại cơ trơn đơn nhất nội tạng của nhiều cơ quan rỗng, là khả năng quay trở lại gần như lực co bóp ban đầu của nó vài giây hoặc vài phút sau khi nó bị kéo dài hoặc rút ngắn.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Thành phần
Cơ trơn có các sợi actin và myosin, tính chất hóa học tương tự như sợi actin và myosin của cơ vân. Tuy nhiên, nó không chứa phức hợp troponin- thành phần kiểm soát co cơ ở cơ vân, vì vậy cơ chế kiểm soát co cơ 2 loại khác nhau. Vấn đề này sẽ được giải thích sau một cách rõ hơn.
Các nghiên cứu chỉ ra rằng, sợi actin và myosin trong cơ trơn hoạt động như cơ chế của cơ vân. Cụ thể là, quá trình này được hoạt hóa bởi ion Ca và adenosine triphosphate (ATP) bị phân giải thành adenosine diphosphate (ADP) để cung cấp năng lượng cho sự co cơ.
Tuy nhiên, có những khác nhau chủ yếu giữa giữa cấu trúc cơ trơn và cơ vân, như khác nhau giữa kích thích-co cơ, cách kênh Ca, duy trì co cơ và năng lượng tiêu hao do co cơ.
Đặc điểm cấu tạo
Sự sắp xếp các sợi actin và myosin là khác nhau giữa cơ trơn khác cơ vân. Dưới kính hiển vi điện tử, một lượng lớn các sợi actin bị bó lại bởi các dense bodies. Một vài dense bodies gắn vào màng tế bào, một số khác nằm trong tế bào cơ. Các dense bodies trên màng tế bào gắn với nhau thông qua các cầu nối protetin nội bào, đây là cơ sở truyền lực co cơ giữa các tế bào.
Xen giữa các sợi actin là các sợi myosin. Sợi myosin có đường kính gấp đôi sợi actin. Dưới kính hiển vi điện tử, số lượng sợi actin gấp 5-10 lần sợi myosin.
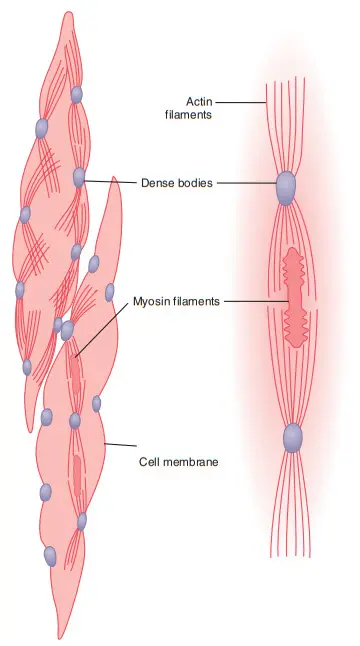
Hình. Cấu trúc vật lý của cơ trơn. Sợi ở phía trên bên trái cho thấy các sợi Actin tỏa ra từ các cơ thể dày đặc. Sợi ở phía dưới bên trái và bên phải chứng minh mối quan hệ của các sợi myosin với các sợi Actin.
Hình bên phải miêu tả một đơn vị cơ trơn. Nhiều sợi actin tỏa ra từ các dense bodies. Tận cùng các sợi actin nằm chồng chéo lên các sợi myosin. Cấu trúc này tương tự như cơ vân nhưng không theo quy luật của cơ vân. Thực tế, các dense bodies đóng vai trò tương tự như các đĩa Z của cơ vân.
Một sự khác biệt nữa là các sợi myosin có các cầu nối “sidepolar” được sắp xếp sao cho mỗi bên gắn với một phía cầu nối. Sự sắp xếp này cho phép các sợi myosin kéo một sợi actin theo một hướng và kéo các sợi actin khác theo hướng ngược lại. Việc này giúp các sợi cơ trơn co ngắn được 80% tổng chiều dài của chúng trong khi ở cơ vân chưa được 30%.
So sánh sự co cơ ở cơ trơn và cơ vân
Trong khi cơ vân co và giãn rất nhanh thì cơ vân co rất chậm, có thể kéo dài nhiều giờ hoặc thậm chí nhiều ngày. Vì vậy, rõ ràng cấu trúc và thành phần giữa 2 loại cơ sẽ khác nhau.
Cầu nối Myosin chậm
Vận tốc của các cầu nối Myosin trong cơ trơn là sự gắn rồi nhả sợi các sợi actin, và lại gắn rồi nhả, chậm nhiều lần so với cơ vân. Thực tế, tần số này ở cơ vân là 1/10 đến 1/300. Tuy nhiên, chính sự gắn chậm này làm tăng lực co cơ ở cơ trơn. Một lý do khác cho sự gắn chậm này là các cầu nối Myosin ở cơ trơn sử dụng ít năng lượng ATP hơn.
Nhu cầu năng lượng để duy trì co cơ thấp
Nhu cầu năng lượng này chỉ bằng 1/10 đến 1/300 so với cơ vân do quá trình gắn và nhả myosin với actin chỉ sử dụng 1 ATP, dù thời gian co cơ trơn kéo dài bao lâu. Điều này có vai trò quan trọng với tổng năng lượng cơ thể bởi các cơ quan như ruột, bàng quang, túi mật… co cơ gần như liên tục.
Thời gian từ lúc kích thích đến khi co và giãn kéo dài
Nhìn chung, khoảng thời gian này từ 0-100 mili giây, co tối đa sau 0.5 giây tiếp, giãn trong 1-2 giây, tổng thời gian 1-3 giây, dài gấp 30 lần so với cơ vân. Tuy nhiên, thời gian kích thích này tùy thuộc từng loại cơ trơn, có thể từ 0.2-30 giây tùy loại. Điều này được giải thích là do các cầu nối Myosin chậm và đáp ứng với ion Ca chậm hơn cơ vân.
Lực co tối đa của cơ trơn mạnh hơn nhiều lần cơ vân
Mặc dù cơ trơn có ít sợi myosin hơn và do các cầu nối Myosin chậm, lực co tối đa của cơ trơn mạnh hơn cơ vân nhiều lần. Ví dụ cùng một cm2, cơ trơn kéo được 4-6kg trong khi cơ vân chỉ kéo được 2-3kg. Nguyên nhân là do khả năng duy trì liên kết giữa sợi actin và myosin.
Cơ chế chốt tạo điều kiện cho các cơn co thắt cơ trơn kéo dài. Một khi cơ trơn đã phát triển co bóp hoàn toàn, lượng kích thích tiếp tục thường có thể giảm xuống thấp hơn nhiều so với mức ban đầu mặc dù cơ vẫn duy trì toàn bộ lực co bóp. Hơn nữa, năng lượng tiêu thụ để duy trì sự co lại thường rất nhỏ, đôi khi chỉ bằng 1/300 năng lượng cần thiết cho sự co cơ xương duy trì tương đương. Cơ chế này được gọi là cơ chế chốt chốt của Wap. Tầm quan trọng của cơ chế chốt là nó có thể duy trì sự co thắt kéo dài trong cơ trơn trong nhiều giờ mà ít sử dụng năng lượng. Ít tín hiệu kích thích tiếp tục được yêu cầu từ các sợi thần kinh hoặc các nguồn nội tiết tố. Căng thẳng-Thư giãn của cơ trơn. Một đặc tính quan trọng khác của cơ trơn, đặc biệt là loại cơ trơn đơn nhất nội tạng của nhiều cơ quan rỗng, là khả năng quay trở lại gần như lực co bóp ban đầu của nó vài giây hoặc vài phút sau khi nó bị kéo dài hoặc rút ngắn. Ví dụ, sự tăng đột ngột về thể tích dịch trong bàng quang tiết niệu, do đó kéo căng cơ trơn trong thành bàng quang, gây ra sự gia tăng áp lực lớn ngay lập tức trong bàng quang. Tuy nhiên, trong 15 giây tiếp theo hoặc lâu hơn, mặc dù tiếp tục kéo dài thành bàng quang, áp lực trở lại gần như chính xác trở lại mức ban đầu. Sau đó, khi âm lượng được tăng thêm bởi một bước khác, hiệu ứng tương tự lại xảy ra. Ngược lại, khi âm lượng giảm đột ngột, áp lực giảm mạnh lúc đầu nhưng sau đó tăng lên trong vài giây hoặc vài phút tới hoặc gần mức ban đầu. Những hiện tượng này được gọi là thư giãn căng thẳng và thư giãn căng thẳng ngược. Tầm quan trọng của chúng là, ngoại trừ trong thời gian ngắn, chúng cho phép một cơ quan rỗng duy trì cùng một áp lực bên trong lòng của nó mặc dù được duy trì, những thay đổi lớn về âm lượng.
Bài viết cùng chuyên mục
Sinh lý nội tiết tuyến thượng thận
Tuyến thượng thận gồm hai tuyến nhỏ úp trên hai thận, mỗi tuyến nặng khoảng 4g. Tuyến thượng thận gồm 2 phần riêng biệt : phần vỏ (80%) và phần tuỷ (20%). Chức năng tuỷ thượng thận, liên quan đến hoạt động hệ giao cảm.
Sự xâm nhập tinh trùng vào trứng: các enzyme thể đỉnh và phản ứng thể đỉnh
Chỉ vài phút sau khi tinh trùng đầu tiên xâm nhập qua lớp màng sáng, các ion calci xâm nhập vào bên trong, tế bào phát động phản ứn g vỏ đổ các chất đặc biệt vào xoang quanh noãn.
Androgen của thượng thận
Androgen thượng thận cũng gây tác dụng nhẹ ở nữ, không chỉ trước tuổi dậy thì mà còn trong suốt cuộc đời, phần lớn sự tăng trưởng của lông mu và nách ở nữ là kết quả hoạt động của các hormon này.
Insulin là hormon liên quan đến sự thừa năng lượng
Carbohydrate dư thừa nếu không thể được dự trữ dưới dạng glycogen sẽ được chuyển thành chất béo dưới sự kích thích của insulin và được dự trữ ở mô mỡ.
Lưu lượng máu mạch vành bình thường
Lưu lượng mạch vành trái giảm trong thời kì tâm thu, khác với lại các mạch khác trong cơ thể. Bởi vì trong thời kì tâm thu, mạch vành bị nén lại mạnh mẽ tho co cơ tim của thất trái.
Tầm quan trọng của trung tâm thưởng phạt của hệ limbic đối với hành vi
Trải nghiệm của con vật qua trải nghiệm cảm giác gây ra trạng thái thưởng hay phạt sẽ được ghi nhớ lại. Điện não đồ cho thấy yếu tố trải nghiệm cảm giác luôn luôn kích thích nhiều vùng trên vỏ não.
Loại dịch trong cơ thể người: đo bằng chỉ thị mầu
Dựa trên nguyên tắc bảo toàn vật chất, có nghĩa là tổng lượng vật chất sau khi hòa vào dịch bằng với tổng lượng vật chất trước khi được bơm vào.
Cơn nhịp nhanh nhĩ: rối loạn nhịp tim
Nhanh nhĩ hay nhanh bộ nối (nút), cả hai đều được gọi là nhịp nhanh trên thất, thường xảy ra ở người trẻ, có thể ở người khỏe mạnh, và những người này thông thường có nguy cơ nhịp nhau sau tuổi vị thành niên.
Các bất thường của áp lực đẩy máu động mạch
Ở người hẹp van động mạch chủ, đường kính của van động mạch chủ khi mở bị hạn chế rõ ràng và áp lực mạch chủ đập cũng giảm rõ bởi vì dòng máu đi ra bị giảm khi qua van bị hẹp đó.
Ngừng tim trong rối loan nhịp tim
Giảm oxy máu khiến cho sợi cơ tim và sợi dẫn truyền mất điện thế màng bình thường và tính dễ kích thích này làm cho nhịp tự động biến mất.
Đo thể tích hô hấp: ghi lại những thay đổi trong phép đo thể tích phổi
Thay đổi của thể tích phổi dưới tình trạng thở khác nhau. Mô tả sự kiện thông khí phổi, không khí trong phổi có thể được chia thành 4 thể tích và 4 dung tích trung bình cho người lớn trẻ.
Chức năng của túi tinh
Tinh dịch có chứa nhiều fructose và nhiều chất dinh dưỡng khác đảm bảo nuôi dưỡng tinh trùng từ lúc xuất tinh đến khi một trong số chúng có thể thụ tinh với trứng.
Sự lắng đọng và hấp thu canxi và phosphate ở xương cân bằng với dịch ngoại bào
Mặc dù cơ chế làm lắng đọng các muối canxi ở osteoid chưa được hiểu đầy đủ, sự kiểm soát của quá trình này dường như phụ thuộc phần lớn vào pyrophosphate, chất làm ức chế tạo thành tinh thể hydroxyapatite và lắng canxi của xương.
Kích thích và dẫn truyền xung động của tim
Nút xoang (còn gọi là nhĩ xoang hay nút SA) phát nhịp trong hệ thống tạo xung nhịp bình thường, theo đường dẫn xung từ nút xoang tới nút nhĩ thất (AV).
Hấp thu ở đại tràng và hình thành phân
Phần lớn hấp thu ở đại tràng xuất hiện ở nửa gần đại tràng, trong khi chức năng phần sau đại tràng chủ yếu là dự trữ phân cho đến một thời điểm thích hợp để bài tiết phân và do đó còn được gọi là đại tràng dự trữ.
Khuếch tán qua màng mao mạch: trao đổi nước và các chất giữa máu và dịch kẽ
Khuếch tán các phân tử nước và chất tan có chuyển động nhiệt di chuyển ngẫu nhiên theo hướng này rồi lại đổi hướng khác. Các chất hòa tan trong lipid khuếch tán trực tiếp qua các màng tế bào ở lớp nội mạc của các mao mạch.
Ức chế đối kháng: cung phản xạ đồi kháng
Phản xạ gấp mạnh hơn này gửi các thông tin ức chế đến chi ban đầu và làm giảm độ co cơ ở chi này, nếu ta loại bỏ kích thích ở chi gấp mạnh hơn, chi ban đầu lại trở về co cơ với cường độ như ban đầu.
Truyền suy nghĩ trí nhớ và thông tin khác giữa hai bán cầu đại não: chức năng thể chai và mép trước trong
Hai bán cầu đại não có khả năng độc lập trong ý thức, trí nhớ, giao tiếp và điều khiển chức năng vận động. Thể chai cần thiết cho hai bán cầu trong các hoạt động phối hợp ở mức tiềm thức nông.
Chức năng và ảnh hưởng của estrogen lên đặc tính sinh dục tiên phát và thứ phát
Trong thời thơ ấu, estrogen chỉ được tiết với một lượng rất nhỏ, nhưng đến giai đoạn dậy thì, lượng estrogen được tiết ra dưới sự kích thích của hormone điều hòa tuyến sinh dục của tuyến yên tăng lên trên 20 lần.
Trạm thần kinh: sự phân kỳ của các tín hiệu đi qua
Sự phân kỳ khuếch đại hiểu đơn giản là các tín hiệu đầu vào lan truyền đến một số lượng nơ-ron lớn hơn khi nó đi qua các cấp nơ-ron liên tiếp trong con đường của nó.
Acid béo với alpha Glycerophosphate để tạo thành Triglycerides
Quá trình tổng hợp triglycerides, chỉ có khoảng 15% năng lượng ban đầu trong đường bị mất đi dưới dạng tạo nhiệt, còn lại 85% được chuyển sang dạng dự trữ triglycerides.
Các đặc trưng của hệ thống điều hòa cơ thể
Một yếu tố nào đó quá tăng hoặc quá giảm, hệ thống điều khiển sẽ thực hiện cơ chế điều hòa đưa nó trở về giá trị bình thường nhờ hàng loạt các biến đổi trong cơ thể, cũng vì thế mà hằng tính nội môi luôn được giữ ổn định.
Điều hòa vận động: vai trò thoi cơ trong các vận động chủ động
Bất cứ khi nào phải thực hiện một động tác yêu cầu độ chính xác và tỉ mỉ, sự kích thích thích hợp từ cấu tạo lưới thân não lên các suốt cơ sẽ giúp duy trì tư thế của các khớp quan trọng, giúp thực hiện các động tác chi tiết một cách khéo léo.
Hormon điều hòa vận động đường tiêu hóa
Phần lớn hormone điều hòa sự bài tiết ở đường tiêu hóa cũng tác động lên sự vận động một số vùng của đường tiêu hóa. Mặc dùng tác dụng vận động thường ít quan trọng hơn tác dụng bài tiết.
Giải phóng năng lượng từ Glucose theo con đường Pentose Phosphate
Con đường Pentose Phosphate có thể cung cấp năng lượng một cách độc lập với tất cả các enzym của chu trình citric acid và do đó là con đường thay thế cho chuyển hóa năng lượng khi có bất thường của enzym xảy ra trong tế bào.
