- Trang chủ
- Sách y học
- Bài giảng miễn dịch
- Một số kháng nguyên quan trọng
Một số kháng nguyên quan trọng
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Hệ kháng nguyên nhóm máu ABO
Hệ này bao gồm 4 nhóm máu khác nhau: A, B, AB và O. Ký hiệu nhóm máu biểu thị kháng nguyên có mặt trên bề mặt hồng cầu. Cơ thể nhóm máu A thì có kháng nguyên A, nhóm B thì có kháng nguyên B, nhóm AB có kháng nguyên A và B và nhóm O thì không có cả A và B trên hồng cầu. Một đặc điểm khác của cơ thể là người máu A có mang kháng thể chống B trong huyết thanh, người máu B thì có huyết thanh chống A, người AB thì huyết thanh không có kháng thể chống hai kháng nguyên này còn nhóm O thì có cả hai loại kháng thể.
Các kháng nguyên thuộc hệ ABO do một locus gen kiểm soát với ba allel A, B, và O, trong đó A và B trội hơn O. Tính đặc hiệu kháng nguyên nhóm máu trong hệ ABO được quyết định bởi sự có mặt của một số gốc “ose” trong phần polysaccharid. Các kháng nguyên này đều có chung một lõi sphingolipid- polysaccharid. Nếu lõi này được gắn thêm gốc fucose thì xuất hiện chất H. Chất này có trên bề mặt hồng cầu của hầu hết các cơ thể và là nền để suất hiện kháng nguyên A và kháng nguyên B. Nếu tại vị trí galactose cuối cùng của chất H có gắn thêm N-acetyl galactosamin thì xuất hiện kháng nguyên A. Còn nếu gắn thêm một galactose nữa thì xuất hiện kháng nguyên B.
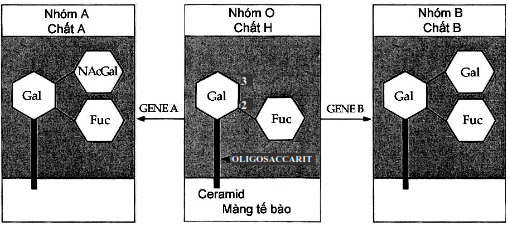
Hình. Sơ đồ cấu tạo kháng nguyên ABO.
Các gen A hoặc B mã hóa cho các enzym transferase gắn Acetylgalactosamin hoặc galactose vào chất H để tạo lên nhóm máu A hoặc B, nếu không có gen A hoặc B thì nhóm máu O.
Về mặt di truyền, tham gia vào sự hình thành kháng nguyên nhóm máu có hai hệ gen, đó là hệ Hh đối với chất H và hệ ABO. Hai hệ này hoạt động độc lập với nhau. Đại đa số chúng ta có gen H, nhưng cũng có người không có (cơ thể đồng hợp tử hh). Những người này dù có gen của hệ ABO nhưng trên hồng cầu vẫn không có kháng nguyên A hoặc B nên được ghi nhận là nhóm O nhưng khi truyền máu O thực sự vào người này (tức đưa chất H vào cho người không có H) thì có thể gây tai biến truyền máu. Nhóm đặc biệt này được gọi là nhóm O Bombay.
Hệ kháng nguyên nhóm máu Rh
Năm 1930, Landsteinner và Wiener đã phát hiện ra nhóm máu Rh. Họ gọi những người có hồng cầu ngưng kết với huyết thanh thỏ kháng hồng cầu của khỉ Rhessus là người Rh+, số người còn lại là Rh-. Nhưng rồi sau này, người ta đã phát hiện ra rằng, kháng nguyên trong hệ Rh không đơn giản như vậy. Hệ Rh có nhiều kháng nguyên, phần lớn có tính sinh miễn dịch yếu và hay gây phản ứng chéo. Chỉ có kháng nguyên D là có tính sinh miễn dịch mạnh. Khi hồng cầu có kháng nguyên D thì được gọi là Rh+ mà không cần để ý đến các kháng nguyên khác trong hệ này. Kháng thể kháng D không xuất hiện tự nhiễm trong máu như các kháng thể của hệ ABO. Vì kháng nguyên Rh phân bố thưa thớt trên hồng cầu nên thường khó gây ngưng kết hồng cầu khi làm phản ứng xác định nhóm máu Rh, vì thế người ta phải dùng đến thử nghiệm Coombs giám tiếp.
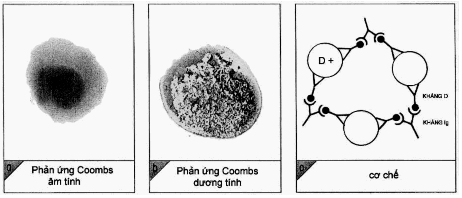
Hình. Phản ứng Coombs gián tiếp
(a). Hình ảnh phản ứng âm tính; (b) Hình ảnh phản ứng dương tính;
(c) Cơ chế phản ứng trong trường hợp bất đồng nhóm máu Rh.
Kháng nguyên vi sinh vật
Kháng nguyên vi sinh vật là loại kháng nguyên rất phức tạp: kháng nguyên vỏ, kháng nguyên thân, kháng nguyên lông, kháng nguyên ngoại tế bào, v.v…
Về kháng nguyên vỏ thì kháng nguyên vỏ của phế cầu thường hay được nghiên cứu nhiều nhất. Kháng nguyên này thuộc loại polysaccaride, có tới 80 typ huyết thanh khác nhau.
Về kháng nguyên thân thì ở các vi khuẩn ruột non, lớp ngoài của vách vi khuẩn có một lipopolisaccharide: phần lipid có tính độc, còn phần polysaccharid thì có tính kháng nguyên và đây chính là kháng nguyên thân (còn gọi là kháng nguyên O). Ở Salmonella có đến hơn 60 kháng nguyên O mà cấu trúc có hai phần: phần nhân cơ bản và phần chuổi ngang. Phần nhân giống nhau ở tất cả các Salmonella. Phần chuổi ngang gồm nhưng tiểu đơn vị oligosaccharide sắp xếp lặp đi lặp lại. Chính các chuổi ngang quyết định tính đặc hiệu của mỗi nhóm Salmonella.
Các kháng nguyên lông của Salmonella có bản chất là protein, còn được gọi là kháng thể H, quyết định tính đặc hiệu của mỗi typ Salmonella. Người ta đã phát hiện Salmonella có hơn 1000 typ huyết thanh khác nhau.
Các kháng nguyên ngoại tế bào như độc tố và các enzym cũng có bản chất protein. Một loại kháng nguyên ngoại tế bào của liên cầu là Streptolysin O thường được dùng trong chẩn đoán huyết thanh. Các vi khuẩn trong bạch hầu, uốn ván có ngoại độc tố gây bệnh và các vaccin phòng các bệnh này đây là độc tố đã được giải độc.
Kháng thể virus có thể ở trên bề mặt hạt virus (capsid) hoặc ở bên trong. Tùy theo tính đặc hiệu, có thể phân biệt các kháng nguyên nhóm, các kháng nguyên typ và kháng nguyên typ phụ. Trong các kháng nguyên nhóm, có thể kể làm ví dụ kháng nguyên nucleoprotein (NPA), là kháng nguyên typ, chẳng hạn trong bệnh bại liệt, cho phép phân biệt ba typ virus bại liệt khác nhau. Trong trường hợp virus cúm, ngoài việc phân biệt ba typ A, B, C, người ta còn phân biệt typ phụ nữa. Các typ phụ của A và B là kết quả của những biến đổi của kháng nguyên bề mặt.
Kháng nguyên hòa hợp mô
Từ lâu, người ta đã biết rằng, việc truyền máu sở dĩ thành công là nhờ, sự phù hợp giữa máu của người cho và máu của người nhận. Trong bài phát biểu khi nhận giải Nobel vào năm 1931, Ladsteiner đã cho rằng có lẽ có “những nhóm giống nhóm máu” liên quan đến việc chấp nhận hoặc thải bỏ những mô ghép khác. Ý tưởng này đã dẫn dắt Gorer đi đến việc xác định một nhóm kháng nguyên ở chuột mà khi nhóm kháng nguyên này giống nhau giữa con cho và con nhận thì mảnh ghép có thời gian sống lâu hơn. Tên gọi kháng nguyên hòa hợp mô đã được dùng để chỉ kháng nguyên tham gia vào việc thải ghép này. Người ta cũng nhận thấy rằng phụ trách mã hóa các kháng nguyên này là một vùng đặc biệt của genom và ta gọi vùng này là Phức hệ hòa hợp mô chủ yếu (Major Histocompatibility Complex, MHC); ở chuột nhắt nó mang tên là vùng H-2, và nằm trên nhiễm sắc thể 17. Những hệ thống tương đương với phức hệ hòa hợp mô chủ yếu ở chuột cũng đã phát hiện được trên tất cả các động vật có vú khác. Ở người, phức hệ hòa hợp mô chủ yếu là cụm gen HLA trên nhiễm sắc thể số 6. Sự tái tổ hợp giữa các cực của phức hệ gen xảy ra khoảng 1% các gia đình, nhờ đó có thể tính được rằng HLA chiếm khoảng 1/3.000 trong toàn bộ genom. Điều này có nghĩa rằng có thể có hàng trăm gen nằm trong cụm HLA này. Mặc dù MHC lần đầu tiên được phát hiện nhờ vai trò của nó trong thải ghép, nhưng ngày nay người ta đã biết rằng, các protein sản phẩm của đoạn gen này đã tham gia vào nhiều công đoạn trong nhận diện miễn dịch, bao gồm tương tác giữa các tế bào lympho khác nhau cũng như các lympho bào với tế bào trình diện kháng nguyên.
Sự sắp xếp các gen MHC
Người ta đã xác định được phức hợp gen MHC mang nhiều gen riêng lẻ; và mặc dù phức hợp có chức năng chung, tương đối giống nhau giữa các loài, nhưng chi tiết về phân bố thì mỗi loài mỗi khác.
Gần đây, người ta đã thiết lập được bản đồ gen hoàn chỉnh cho MHC ở chuột nhắt. Nhờ đó mà chúng ta biết được rằng locus nào thì chịu trách nhiệm mã hóa cho đoạn polypeptid nào. Chỉ còn điều vướng mắc là cách thức các polypeptid và protein thực hiện chức năng của mình như thế nào cho phù hợp với quy định của MHC mã hóa tìm ra được là nhờ kỹ thuật miễn dịch học (dùng kháng thể đơn clôn), do đó mà người ta thường gọi các protein này là kháng nguyên MHC.
Các protein của MHC gồm có 3 lớp khác nhau về cả cấu trúc lẫn chức năng:
Protein lớp I (tức kháng nguyên MHC lớp I) gồm có hai polypeptid. Peptid lớn được mã hóa bởi MHC, peptid này liên kết với một polypeptid thứ hai là β2- microglobulin được mã hóa bởi một gen ngoài đoạn MHC.
Các protein lớp II (tức kháng nguyên MHC lớp II) gồm có hai peptid không đồng hóa trị với nhau có tên là chuỗi α và chuỗi β, cả hai đều là sản phẩm của MHC.
Các protein lớp III (tức kháng nguyên MHC lớp II) là các thành phần bổ thể được mã hóa bởi MHC.
Ngoài ra còn có những locus phụ nằm ngoài phức hợp H-2 (phức hợp TLA) tham gia mã hóa cho protein lớp I và bởi vì những protein này cũng có chức năng miễn dịch nên cũng được đưa vào bản đồ gen ở.
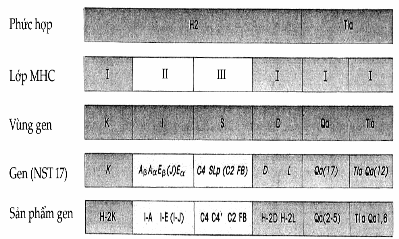
Hình. Sơ đồ vị trí các gen MHC
Có 17 gen Qa trong vùng Qa và 12 gen trong vùng Tla mặc dù đa số chúng ở dạng nghỉ.
Ở người, những vùng chính là D, B, C, và A. Các vùng A và B ở người là vùng có chức năng tương đương với vùng K và D ở chuột, điều đó có nghĩa rằng những sản phẩm của chúng tác động với tư cách là những phân tử nhận diện trên bề mặt tế bào, và tế bào T gây độc sẽ nhận diện những phân tử này khi thực hiện chức năng của nó. Vùng D của người tương đương với vùng I của chuột và sản phẩm của chúng (hai chuỗi protein α và β) tham gia vào quá trình hợp tác của tế bào giữa tế bào của hệ miễn dịch. Các sản phẩm vùng I của chuột được gọi là kháng nguyên Ia. Tương đương với kháng nguyên DR, DP, và DQ của người. Người ta nhận thấy rằng, cường độ của đáp ứng miễn dịch trên chuột được xác định một phần bởi các gen đáp ứng miễn dịch.
Phân bố của kháng nguyên MHC
Ở người, tất cả các tế bào có nhân đều nhất thiết phải mang các kháng nguyên của vùng A, B, C, nhưng còn kháng nguyên được mã hóa bởi vùng D thì chỉ phân bố hạn chế ở những tế bào tương đương với tế bào chuột có mang kháng nguyên I. Kháng nguyên lớp I chỉ có trên tế bào B, đại thực bào, tế bào mono và có lẽ cả tế bào biểu mô. Ngoài ra, một số kháng nguyên lớp I xuất hiện cả trên tế bào T ức chế và kháng nguyên lớp II thì có trên tế bào T được hoạt hóa.

Bảng. Phân bố của kháng nguyên MHC ở các mô.
Cấu trúc của kháng nguyên MHC
Những công trình gần đây nghiên cứu các kháng nguyên MHC riêng biệt đã giúp làm sáng tỏ cấu trúc của chúng. Với tư cách là phân tử tham gia vào tương tác tế bào, kháng nguyên MHC thường được tìm thấy trên màng tế bào. Chúng là những phân tử glycoprtein cắm vào màng tế bào, do đó để nghiên cứu được chúng ta thường phải tách rời chúng khởi màng bằng hai cách:
Hoặc là dùng các chất tẩy (detergent) để giải phóng chúng khỏi màng.
Hoặc là dùng enzym papain để cắt phần MHC nằm thòi ra khỏi bề dày của màng. Khi phân tử này được cắt bằng enzym thì chúng có khác với kháng nguyên MHC nguyên vẹn bởi vì chúng đã mất phần cắm sâu vào màng tế bào. Người ta làm tinh khiết kháng nguyên hòa tan bằng các kỹ thuật hóa sinh kinh điển như sắc ký ái tính, dùng lectin hoặc kháng huyết thanh. Người ta nhận thấy rằng tinh luyện kháng nguyên MHC bằng sắc ký ái tính trên cột kháng thể đơn clôn có nhiều ưu điểm hơn các kỹ thuật hóa sinh khác.
Mỗi kháng nguyên lớp I bao gồm một polypeptid được gắn đường có trọng lượng phân tử khoảng 45.000 liên kết không đồng hóa trị với một peptid không có đường (tức β2 -microglobulin, có trọng lượng phân tử khoảng 12.000). Dù β2 - microglobulin không tham gia vào bề mặt kháng nguyên của phân tử HLA, nhưng nó cần cho quá trình thể hiện của lớp I. Nếu một cá thể nào đó thiếu bẩm sinh β2 - microglobulin thì quyết định kháng nguyên của lớp I không thể hiện được.
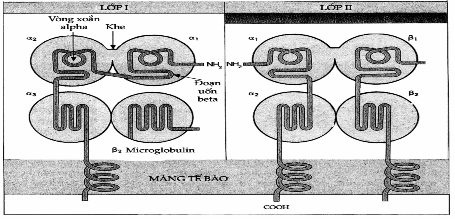
Hình. Sơ đồ minh họa cấu trúc các phân tử MHC lớp I và II.
Sơ đồ cho thấy các phân tử MHC luôn có cấu trúc cắm sâu vào màng tế bào.
Cấu trúc của lớp II ít được biết rõ hơn lớp I, chúng gồm hai chuỗi poypeptid khác nhau α và β gắn với nhau bởi lực nối không đồng hóa trị, cả hai chuỗi đều cắm vào màng tế bào. Chuäùi ngắn mang các kháng nguyên allotyp, và hai chuäùi đều có mang các đơn vị đường.
Người ta biết nhiều về cấu trúc của vùng H-2I của chuột hơn vùng tương đương với vùng này ở người là vùng HLA-D. Trên chuột, người ta đã phát hiện thêm những tiểu vùng của H-2I, đó là tiểu vùng I-A và I-E.
Khảo sát riêng từng vùng peptid và của kháng nguyên I-E ở chuột cho thấy rằng, đa số các biến đổi cấu trúc chỉ xảy ra ở chuỗi β (trường hợp kháng nguyên HLA-D cũng vậy).
Chức năng của kháng nguyên MHC
Các kháng nguyên MHC rất cần thiết cho các quá trình nhận diện miễn dịch. Các kháng nguyên MHC khác nhau được nhận diện bởi những loại tế bào T khác nhau. Những tế bào T gây độc tham gia vào việc nhận diện và loại bỏ các tế bào nhiễm virus và mô ghép lạ có khả năng nhận diện các phân tử H-2K và H-2D (tương đương HLA-A và HLA-B ở người) trên các tế bào lạ, và khi hợp tác với tế bào T giúp đỡ thì sẽ gây phá hủy tế bào lạ.
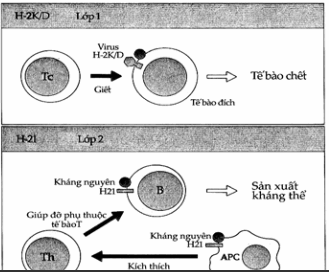
Hình. Tóm tắt các chức năng của MHC ở chuột.
Tế bào T gây độc nhận diện kháng nguyên lạ kết hợp với H- 2K/D và tiêu diệt tế bào đích.
Tế bào T giúp đỡ nhận diện kháng nguyên kết hợp với H-2I trên tế bào trình diện kháng nguyên.
Tế bào Ts ức chế đặc hiệu hoạt động của đại thực bào, tế bào B và Th. Đây là các ức chế phụ thuộc H- 2IJ.
Đa số tế bào nhiễm virus thể hiện kháng nguyên virus trên bề mặt của chúng, và những kháng nguyên này được nhận diện bởi tế bào T gây độc. Đồng thời người ta cũng biết được rằng tế bào T gây độc chỉ nhận diện được kháng nguyên virus khi có phối hợp với việc nhận diện kháng nguyên K-2K và -D có mặt trên tế bào bị nhiễm. Bằng cách này tế bào T gây độc có thể giết chết tế bào nhiễm.
Như vậy, phân tử H-2 trên bề mặt tế bào nhiễm virus tác động như một dấu hiệu hướng dẫn đưa tế bào T gây độc đến tiếp cận bào đích, và giúp nó để phân biệt tế bào đích với các tế bào khác mang kháng nguyên virus.
Người ta vẫn chưa biết rõ rằng tế bào gây độc đã nhận diện cùng một lúc tính đặc hiệu virus và kháng nguyên MHC tế bào đích như thế nào. Có hai giả thuyết.
Tế bào gây độc đã nhận diện độc lập hai kháng nguyên này.
Sự kết hợp của kháng nguyên virus và kháng nguyên MHC trên bề mặt tế bào tế bào đã làm biến đổi hình dạng cấu trúc của phân tử MHC, do đó mà nó nhận diện như một thành phần bản thân đã bị biến đổi.
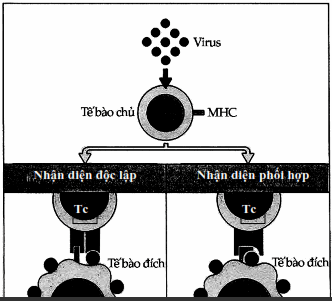
Hình. Hai giả thuyết về việc tế bào Tc nhận diện kháng nguyên virus và MHC trên bề mặt tế bào đích.
Tế bào Tc có thể nhận diện độc lập các phân tử kháng nguyên virus và MHC (giả thuyết 1) hoặc nhận diện chung phức hợp tạo nên bởi kháng nguyên virus và MHC (giả thuyết 2).
Nhiều chức năng của MHC khác sẽ được đề cập cụ thể hơn, chúng ta nghiên cứu các bộ phận của hệ thống miễn dịch vì MHC tham gia vào hết các quá trình nhận diện miễn dịch. Những hiểu biết gần đây cho thấy rằng, MHC và phân tử khác tham gia và nhận diện miễn dịch giống nhau ở một phần cấu trúc, và điều này có lẽ có liên quan về mặt tiến hóa với những phân tử nhận diện đầu tiên trong thời kỳ bào thai.
Có một điều chúng ta cần lưu ý là có rất nhiều thay đổi allotyp xảy ra cho các kháng nguyên MHC. Sự xuất hiện của những tính đặc hiệu mới của MHC có thể thấy ở những loài khác nhau và có liên quan với nhau. Nguyên nhân của sự thay đổi lớn này chưa được rõ. Vì tính chất rất đặc thù của từng cá thể sinh vật mà trong một quần thể không có yếu tố bệnh nguyên nào có thể gây biến đổi giống nhau cho toàn bộ các cá thể ở mức độ nhận diện kháng nguyên. Tuy vậy, chúng ta cần biết rằng trong một quần thể có thể có hiện tượng một cá thể mang một kháng nguyên MHC đặc biệt nào đó thì có tính dễ mắc một bệnh tương ứng hoặc không mắc một bệnh tương ứng so với những người khác. Đó là đặc điểm di truyền của MHC.
Bài viết cùng chuyên mục
Định lượng immunoglobulin và các protein đặc hiệu khác
Kỹ thuật thường được dùng phổ biến nhất, là miễn dịch kết tủa, tủa miễn dịch được hình thành khi kháng nguyên, và kháng thể, kết tủa tương ứng.
Đánh giá tế bào trung tính và tế bào mono
Thực bào là chức năng ăn vật lạ của một tế bào nào đó, khả năng ăn này có thể xác định được bằng cách ủ tế bào thực bào, với các hạt trơ như hạt latex, hoặc vi khuẩn.
Đại cương bổ thể
Các protein của hệ thống bổ thể tạo thành hai chuỗi enzym, mà người ta gọi là con đường cổ điển, và con đường không cổ điển, để tạo nên hai cách phân cách hai phân tử C3
Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Một phân tử IgM pentamer, kết hợp với kháng nguyên, là có thể cố định bổ thể, nhưng đối với IgG, thì phải có phân tử IgG được gắn với kháng nguyên ở vị trí gần nhau.
Con đường hoạt hóa bổ thể không cổ điển (alternative pathway)
Không nhất thiết phải có phản ứng kháng nguyên, kháng thể, nên phản ứng có thể xảy ra tức thì, và cơ thể được bảo vệ theo cơ chế không đặc hiệu.
Sự kết hợp kháng nguyên kháng thể
Khả năng trung hòa độc tố, và vi sinh vật của kháng thể, luôn phụ thuộc vào sự gắn kết chặt chẽ của chúng vào kháng thể.
Kỹ thuật DNA tái tổ hợp và miễn dịch lâm sàng
Kỹ thuật DNA tái tổ hợp, dựa vào việc sử dụng plasmid như các phương tiện để truyền các đoạn DNA lạ, ví dụ gen người.
Chẩn đoán và tiên lượng bệnh miễn dịch
Tự kháng thể cón có giá trị tiên lượng, trường hợp một đứa trẻ, có anh chị em mắc bệnh đái đường phụ thuộc insulin, nó có chung HLA với anh chị.
Đại cương thiếu hụt miễn dịch
Một đứa trẻ, hay một người lớn, sẽ bị nghi ngờ là thiếu hụt miễn dịch, khi trên cơ thể xuất hiện những nhiễm trùng lặp đi lặp lại, tồn tại kéo dài, trầm trọng hoặc bất thường.
Đại cương các tế bào chủ yếu của hệ thống miễn dịch
Tế bào tham gia vào đáp ứng miễn dịch thu được, bao gồm các lymphô bào đặc hiệu kháng nguyên, tế bào trình diện kháng nguyên.
Đại cương miễn dịch chống vi sinh vật
Đối với nhiễm trùng, một cân bằng được duy trì giữa sức chống đỡ của cơ thể, và khả năng của vi sinh vật cố gắng để vượt qua sức chống đỡ đó.
Sự sắp xếp các bệnh tự miễn thường gặp ở người
Những cơ quan đích thường gặp, trong bệnh đặc hiệu cơ quan, là tuyến giáp, tuyến thượng thận, dạ dày và tuyến tụy.
Sản xuất kháng huyết thanh cho các phòng thí nghiệm miễn dịch lâm sàng
Hỗn dịch tế bào lách của con vật được gây mẫn cảm, có chứa nhiều tế bào B chịu trách nhiệm sản xuất nhiều kháng thể, chống nhiều epitope khác nhau.
Cấu trúc phân tử của kháng thể
Các glycoprotein huyết tương, hoặc huyết thanh trước đây thường được phân chia theo tính chất hòa tan, của chúng thành albumin và globulin.
Miễn dịch chống ký sinh trùng
Bệnh nhân chống chọi với nhiễm ký sinh trùng đơn bào, bằng những phản ứng tương tự như trong nhiễm vi khuẩn, có một số đơn bào có cơ chế tồn tại độc đáo.
Các tính chất của kháng nguyên
Một kháng nguyên protein phức tạp, có thể nhiều quyết định kháng nguyên khác nhau, do đó mà nó có thể kích thích tạo ra nhiều loại kháng thể khác nhau.
Các cytokin trung gian và điều hòa miễn dịch bẩm sinh
Có 2 loại thụ thể của TNF, loại có trọng lượng phân tử 55 kD có tên là THF RI, và loại có trọng lượng phân tử 75 kD có tên là TNF RII.
Cơ chế bệnh sinh bệnh miễn dịch
Tế bào B tự phản ứng, tế bào T hiệu quả và tự kháng nguyên bình thường, vẫn có mặt trong cơ thể, nhưng không được khởi động.
Khảo sát định tính immunoglobulin
Trong trường hợp không có bất thường chuỗi nặng, kháng huyết thanh chuỗi nhẹ tự do, tức không phản ứng với chuỗi nhẹ cố định vào chuỗi nặng.
Miễn dịch chống virus
Virus Epsptein-Barr dùng thụ thể của C3b tức CR2, còn HIV thì lại dùng thụ thể CD4 để làm nơi xâm nhập vaof tế bào đích, trong hệ thống miễn dịch.
Kháng thể đối với kháng nguyên ngoại sinh
Thử nghiệm kích thích provocation test, tức thử nghiệm kích thích niêm mạc mũi hoặc niêm mạc phế quản bằng kháng nguyên, là một thử nghiệm khá phổ biến.
Quá mẫn miễn dịch typ III
Kháng thể và kháng nguyên tạo thành phức hợp, phức hợp này hoạt hóa bổ thể đồng thời tác động gây giải phóng các amin hoạt mạch, làm tăng tính thấm thành mạch.
Quá mẫn miễn dịch typ IV (Quá mẫn muộn)
Cần luôn nhớ rằng, tổn thương quá mẫn, là hậu quả của phản ứng quá mức, giữa kháng nguyên, với cơ chế miễn dịch tế bào vẫn còn bình thường
Tế bào lymphô hệ miễn dịch
Tế bào lymphô bao gồm nhiều tiểu quần thể khác nhau, khác biệt về chức năng, về sản phẩm protein nhưng không phân biệt được về hình thái.
Các cytokin trung gian và điều hòa miễn dịch thu được
Mặc dù lúc đầu người ta phát hiện ra IL 2, như là một yếu tố phát triển tế bào T, nhưng thật ra IL 2 có nhiều chức năng trong đáp ứng miễn dịch thu được.
