- Trang chủ
- Sách y học
- Bài giảng miễn dịch
- Miễn dịch chống virus
Miễn dịch chống virus
Virus Epsptein-Barr dùng thụ thể của C3b tức CR2, còn HIV thì lại dùng thụ thể CD4 để làm nơi xâm nhập vaof tế bào đích, trong hệ thống miễn dịch.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Virus và đáp ứng miễn dịch
Virus có những tính chất độc đáo riêng:
(1) chúng có thể xâm nhập vào mô mà không gây ra một đáp ứng viêm.
(2) chúng có thể nhân lên trong tế bào trong suốt đời sống cơ thể chủ mà không gây ra tổn thương tế bào.
(3) đôi khi chúng cản trở một số chức năng đặc biệt của tế bào mà không gây biểu hiện ra ngoài.
(4) cũng có khi virus gây tổn thương mô hoặc cản trở sự phát triển tế bào và rồi biến mất hoàn toàn khỏi cơ thể.
Gần đây, người ta phát hiện được rằng một số bệnh trước đây không rõ nguyên nhân như viêm não xơ hóa bán cấp, bệnh não chất trắng đa tiêu điểm tiến triển (progressive multifocal leukoencephalopathy) bệnh Creuzfeld - Jacob, bệnh Curu lại là những bệnh có liên quan đến virus hoặc những vật thể giống virus. Biểu hiện lâm sàng của bệnh virus rất đa dạng.
Nhóm virus herpes bao gồm ít nhất 60 virus, trong đó có 5 con rất hay gây bệnh cho người: Herpes simplex (HSV) typ 1, HSV typ 2, Varicella zoster (VZV), cytomegalovirus (CMV) và Epstein-Barr (EBV). Có hình ảnh bệnh lý chung cho tất cả các virus nhóm herpes ở người, đó là: Một, để truyền được người này sang người khác phải có sự tiếp xúc gần gũi trực tiếp, trừ việc truyền máu và ghép cơ quan là đường truyền hiệu quả nhất của CMV. Hai, sau lần nhiễm đầu tiên virus herpes sẽ tồn tại trong cơ thể suốt đời.
Để hạn chế sự lan tỏa của virus và phòng ngæìa tái nhiễm hệ thống miễn dịch phải có khả năng chặn đứng sự xâm nhập của các hạt virion và tế bào cũng như loại bỏ các tế bào bị nhiễm để giảm nơi cư trú của virus. Như vậy, các phản ứng miễn dịch có hai loại: Một loại để tác động lên các hạt virus và một loại tác động lên tế bào nhiễm. Một cách tổng quát, đáp ứng miễn dịch chống virion có xu hướng trội về thể dịch còn đáp ứng tế bào thì tác dụng lên tế bào nhiễm virus. Cơ chế thể dịch chủ yếu là trung hòa, nhưng phản ứng thực bào phụ thuộc bổ thể và phản ứng ly giải phụ thuộc bổ thể cũng có thể xảy ra.
Trung hòa virus ngăn cản sự tiếp cận của chúng đến các tế bào đích. Đây là chức năng của kháng thể IgG trong dịch ngoại bào và của IgA trên bề mặt niêm mạc. Chúng ta cần nhớ rằng, chỉ những kháng thể chống lại các thành phần chịu trách nhiệm về khả năng tiếp cận mới có tính trung hòa: Sự tạo ra kháng thể có độ đặc hiệu chính xác là nguyên tắc cơ bản để sản xuất vacxin virus. Những kháng thể chống lại những kháng nguyên không cần thiết không chỉ không có tác dụng bảo vệ mà còn tạo điều kiện để hình thành phức hợp miễn dịch.
Mặt dù chỉ cần kháng thể IgG là đủ để trung hòa hầu hết virus, nhưng sự hoạt hóa bổ thể tỏ ra cũng rất có ích trong việc làm tăng cường khả năng loại trừ virus. Sự ly giải virus cũng có thể thực hiện chỉ nhờ vào bổ thể mà không cần có kháng thể. Một số virus như EBVcó thể gắn với C1và hoạt hóa bổ thể theo đường cổ điển để cuối cùng là hạt virion bị ly giải.
Miễn dịch qua trung gian tế bào liên quan tế bào bị nhiễm virus hơn là virus tự do. Lymphô T nhận diện virus trong sự phối hợp với các glycoprotein của phức hệ hòa hợp mô chủ yếu (MHC). Tế bào T gây độc sẽ ly giải tế bào đã bị virus đột nhập hoặc làm thay đổi kháng nguyên bề mặt. Như vậy, miễn dịch tế bào (tế bào T) chịu trách nhiệm trong quá trình hồi phục sau nhiễm virus, chứ không phải trong giai đoạn cấp tính của bệnh.
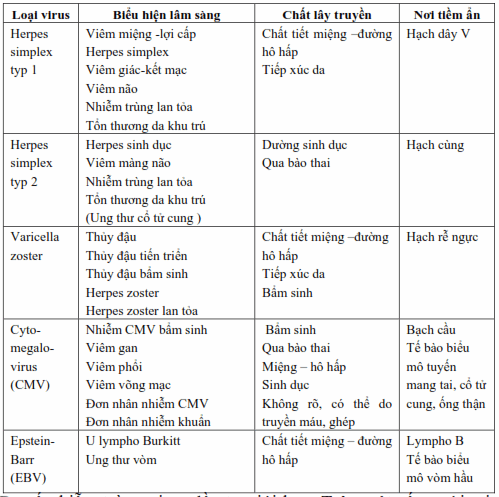
Bảng. Hình ảnh lâm sàng của nhiễm virus herpes.
Đa số nhiễm trùng virus đều tự giới hạn. Trên một số người, virus có thể tạo ra triệu chứng lâm sàng trong lúc đó trên một số người khác thì bệnh không bao giờ vượt quá giai đoạn tiền lâm sàng. Sự hồi phục sau nhiễm trùng virus cấp thường để lại tính miễn dịch lâu dài và ít khi cơ thể bị tấn công lần hai bởi cùng loại virus đó.
Tác động trực tiếp của virus
Hướng tính của virus (viral tropism) là một yếu tố cơ bản để xác định tầm quan trọng lâm sàng của nhiễm trùng virus, là một nhiễm trùng phụ thuộc không những vào số lượng tế bào bị phá hủy còn phụ thuộc vào chức năng của những tế bào đó. Sự phá hủy một số lượng nhỏ tế bào coï chức năng biệt hóa cao như dẫn truyền thần kinh hoặc điều hòa miễn dịch cũng có thể gây nguy hiểm cho tính mạng bệnh nhân. Ngược lại, sự phá hủy một lượng lớn những tế bào ít biệt hóa như tế bào biểu mô chẳng hạn nhưng hậu quả lại ít trầm trọng hơn nhiều.
Càng ngày người ta càng hiểu biết nhiều hơn về các thụ thể virus và tương tác giữa thụ thể này với virus. Virus Epsptein-Barr dùng thụ thể của C3b (tức CR2), còn HIV thì lại dùng thụ thể CD4 để làm nơi xâm nhập vaìo tế bào đích trong hệ thống miễn dịch.
Một khi vaìo trong tế bào, virus có thể giết tế bào bằng nhiều cách. Một số virus như polyovirus, adenovirus và các sản phẩm của chúng có thể ức chế các enzym cần thiết cho sự nhân lên hoặc chuyển hóa tế bào. Một số virus khác có thể phá hủy cấu trúc nội bào như tiêu thể chẳng hạn làm giải phóng ra các enzym độc hại làm chết tế bào. Trong một số trường hợp, protein của virus gắn với màng tế bào làm thay đổi tính chất của nó: Ví dụ như virus sởi có hoạt tính gây hòa màng làm cho các tế bào kết hợp với nhau thành những hợp bào (syncytia).
Một số virus có thể làm thay đổi chức năng đã chuyển hóa của tế bào mà không giết chết chúng. Những tế bào này thường là tế bào của hệ miễn dịch, thần kinh trung æång hoặc hệ nội tiết. Ví dụ các virus sởi, cúm, CMV thường nhiễm vào tế bào lympho người và biến đổi chức năng của chúng. Trong suốt thời kỳ nhiễm trùng, các virus biến tướng được sinh sản một cách chọn lọc trong lách và có thể ức chế sự hình thành các tế bào T gây độc. Hậu quả là virus không bị loại bỏ khỏi cơ thể mà sống suốt đời với ký chủ.
Lẩn tránh của virus (cơ chế tồn tại của virus trong cơ thể)
Virus thường tạo ra nhiều cơ chế khác nhau để lẩn tránh hoặc ngăn cản tác động của hệ miễn dịch.
Thay đổi tính kháng nguyên là cách thấy rõ nhất đối với virus cúm A, một loại virus RNA được bỏ bởi một vỏ lipid có gắn với hai loại protein: hemagglutinin và neuraminidase. Đa số kháng thể trung hòa tác động lên hai quyết định kháng nguyên này. Virus có thể lẩn tránh tác động của kháng thể bằng cách thay đổi cấu trúc của hemagglutinin: Thay đổi dần tính kháng nguyên (antigenic drift) hoặc đột biến tính kháng nguyên (antigenic shift).
Thay đổi dần tính kháng nguyên là sự thay đổi từng phần nhỏ cấu trúc kháng nguyên khi virus truyền từ các thể này sang cá thể khác bằng cách gây đột biến điểm trên bề mặt kháng nguyên của hemagglutinin. Thay đổi này có lẽ chịu trách nhiệm về các dịch cúm nhỏ vào mùa đông.
Thay đổi tính kháng nguyên là sự thay đổi đột ngột toàn bộ cấu trúc của hemagglutinin. Người ta đã quan sát thấy 3 lần thay đổi kháng nguyên kiểu này vào vụ đại dịch cúm năm 1918, dịch cúm châu Á năm 1957, và dịch cúm Hồng Kông năm 1968.
Nếu đáp ứng miễn dịch không loại trừ được hoàn toàn virus thì sẽ xuất hiện một tình trạng nhiễm trùng nhẹ với sự tồn tại dai dẳng của một số virus trong cơ thể. Ví dụ, viêm gan B có thể tồn tại nhiều tháng hoặc nhiều năm và gan liên tục mang virus.
Ngoài ra, virus cũng có thể tạo ra tçnh trạng tiềm tàng nếu genom virus tồn tại mãi trong tế bào chủ mà không thể hiện tính kháng nguyên virus. Tất cả các virus herpes người đều có thể tồn tại tiềm ẩn, thỉnh thoảng có những đợt hoạt động và nhân lên. Khi sự cân bằng giữa virus và cơ thể chủ bị phá vỡ do nhiễm trùng, rối loạn chuyển hóa, tuổi già , hoặc ức chế miễn dịch thì virus được hoạt hóa và sau đó có thể gây ra bệnh. Thường thường mỗi virus có nơi tiềm ẩn riêng của nó: Ví dụ virus herpes simplex tiềm ẩn ở hạch dây V, Varicella zoster tiềm ẩn ở hạch rễ thần kinh ngực .
Sự chuyển dạng tế bào chủ có thể xảy ra do tác động của một số virus có khả năng gây bệnh ung thư. Hầu hết các virus loại này tồn tại tiềm ẩn, ví dụ, HTLV-I gây bệnh bạch cầu tế bào T ở người lớn, virus viêm gan B gây bệnh ung thư tế bào gan và EBV có thể gây ung thư vòm họng hoặc u lymphô Burkitt.
|
BỆNH ÁC TÍNH |
VIRUS |
|
Một số bệnh bạch cầu tế bào T U lymphô Burkitt |
Virus bệnh bạch cầu tế bào T ở người Virus Epstein-Barr |
Bảng. Virus và các bệnh ác tính liên quan.
Một số virus có thể ngăn cản đáp ứng miễn dịch bằng cách ức chế hoặc nhiễm vào tế bào miễn dịch.
Ví dụ, nhiễm trùng CMV tiên phát gây bệnh điển hình ở người trẻ. Đáp ứng tạo kháng thể xảy ra nhanh, có thể thấy được, nhưng đáp ứng miễn dịch tế bào thì lại bị ức chế: Trong nhiều tháng có thể không thấy có đáp ứng tế bào T đặc hiệu với CMV. Ngoài ra, trên thực nghiệm người ta còn thấy rằng miễn dịch tế bào đối với các lần thử thách kháng nguyên trước không xuất hiện, có khi đến cả năm trời. Trong thời gian này, bệnh nhân có thể bị nhiễm trùng vi khuẩn làûp đi làûp lại nhiều lần. Hiệu quả gây ức chế miễn dịch này cũng thấy ngay cả những người đã có ức chế miễn dịch từ trước, ví dụ như người nhận mảnh ghép chẳng hạn.
Virus sởi có khả năng nhân lên trong tế bào T dẫn đến kết quả là gây giảm miễn dịch tế bào. Trước đây, khi chưa điều trị được bệnh lao, bệnh nhân mắc bệnh sởi cấp dễ dàng dẫn đến mắc lao kê.
Ví dụ điển hình nhất cho hiện tượng này là hội chứng suy giảm miễn dịch mắc phải (AIDS) do HIV gây ra. Virus này đã chọn lọc tế bào lymphô T CD4+ để tiêu diệt. Trình trạng ức chế miễn dịch sau đó dẫn đến sự xuất hiện của các nhiễm trùng cơ hội lan tỏa và trầm trọng hoặc các bệnh cảnh ung thư đặc biệt cho bệnh này.
Tổn thương do đáp ứng miễn dịch chống virus
Mặc dù phản ứng miễn dịch nói chung là có lợi. Thỉnh thoảng chúng cũng gây ra tổn thương mô khó phân biệt với tổn thương do chính virus gây ra.
|
TẾ BÀO |
VIRUS |
HẬU QUẢ |
|
Lymphô B
|
Epstein- Barr
HTLV- I
Dengue Lassa Marburg-Ebola |
Chuyển dạng và hoạt hóa tế bào B đa clôn
Nhân lên trong tế bào T hoạt hóa U lymphô hoặc bệnh hạch cầu tế bào T AIDS Sốt
|
Bảng. Một số hậu quả sau nhiễm virus của tế bào hệ miễn dịch.
Trong thời kỳ hồi phục sau nhiễm trùng virus (ví dụ đơn nhân nhiễm khuẩn hoặc viêm gan B), trên một số bệnh nhân người ta thấy xuất hiện các tự kháng thể lưu động do tế bào B vẫn còn khả năng đáp ứng với kháng nguyên bản thân khi chúng được trình diện một cách thích hợp cho tế bào T. Dung nạp miễn dịch bình thường đối với kháng nguyên tự thân do tế bào T ức chế đảm trách là chủ yếu, nhiễm trùng virus đã phá vỡ sự dung nạp này bằng hai cách:
(1) những virus như EBV là những tác nhân hoạt hóa tế bào B đa clôn, và
(2) virus có thể gắn lên kháng nguyên bản thân để làm cho chúng trở thành kháng nguyên mới. Kháng thể đối với những kháng nguyên mới này sẽ tác động lên cả các mô tự thân bình thường lẫn các tế bào đã nhiễm virus. Sự tồn tại lâu dài của nhiễm trùng virus có thể dẫn đến bệnh tự miễn trên một cá thể thích hợp. Một số ví dụ hình thành bệnh gan tự miễn mạn tính trên một số bệnh nhân bị viêm gan siêu vi B.
Một số virus lại có khả năng kích thích tạo những kháng thể không thích hợp làm tăng thêm tổn thương mà virus đã gây cho cơ thể chủ. Ví dụ, virus Dengue có thể nhiễm vào đại thực bào qua đường thụ thể Fc, và khả năng của nó xâm nhập vào tế bào đích sẽ được tăng cường nếu nó gắn với kháng thể IgG. Như vậy, nhiễm trùng lần thứ hai bởi một týp huyết thanh khác sẽ được làm dễ bởi kháng thể do týp huyết thanh trước tạo ra.
Sự tấn công của kháng thể vào những tế bào nhiễm virus có thể gây ra hiệu quả bất lợi. Kháng thể kháng virus hoặc phức hợp miễn dịch tạo nên do virus và kháng thể có thể ngăn cản không cho tế bào lympho nhận diện hoặc phản ứng với kháng nguyên virus, do đó ức chế đáp ứng miễn dịch tế bào; đó là trường hợp của viêm não xơ hóa bán cấp. Trong trường hợp này hình ảnh bệnh lý cho thấy có sự mất myelin thay thế bằng xơ hóa liên tục dẫn đến các rối loạn thần kinh trầm trọng. Khoảng một næía trong số bệnh nhân này đã từng bị mắc sởi vào hai năm tuổi đầu tiên. Trong khi đó trong nhân dân thì tỉ lệ người mắc sởi chiếm tỉ lệ cao nhất vào lứa tuổi từ 5 đến 9. Sự tiếp xúc với virus sởi vào thời điểm sớm của đời sống có lẽ đã giúp cho virus dãù dàng tồn tại trong não dưới dạng virus hoàn chỉnh. Bởi vì cơ thể đã sản xuất một lượng kháng thể kháng sởi tương đối cao, phức hợp miễn dịch tạo bởi virus và kháng thể đã ức chế phản ứng miễn dịch tế bào.
Người ta cho rằng nhiều bệnh viêm mạn tính ở người như viêm cầu thận mạn có liên quan đến sự lắng đọng của phức hợp miễn dịch bởi vì chúng rất giống với các mô hình virus thực nghiệm. Tuy vậy, trong đa số các trường hợp, virus bệnh nguyên tương đối khó xác định.
Một ví dụ kinh điển cho các tổn thương gây ra do tế bào là viêm màng não - màng mạch lymphô ở chuột. Chuột được cho nhiễm virus trong thời kỳ sơ sinh, virus nhân lên nhanh chóng trong nhiều mô, kể cả thần kinh trung ương. Tuy nhiên, điều này không gây bệnh nặng. Ngược lại, nếu tiêm virus vào não của chuột trưởng thành thì có thể gây ra viêm não-màng não nặng dẫn đến tử vong; tổn thương não ở đây có thể hạn chế bằng cách làm rối loạn miễn dịch tế bào và đồng thời có thể duy trì trở lại tổn thương sau đó bằng cách tiêm cho con vật tế bào T đã mẫn cảm từ con vật khác. Các lympho T gây độc từ con vật mẫn cảm nguyên virus trên bề mặt. Như vậy, chúng ta hoàn toàn có lý do để nghi ngờ rằng cơ chế miễn dịch tế bào đã có vai trò trong việc gây viêm não trong nhiều bệnh cảnh nhiễm trùng virus ở người.
Miễn dịch tế bào có lẽ còn chịu nhiều trách nhiệm trong việc gây ra các loại ban đỏ đặc trưng thường gặp trong nhiễm trùng virus ở trẻ con, như trong sởi chẳng hạn. Các tiêu điểm nhỏ virus ở da có thể kích thích tạo một
phản ứng quá mẫn muộn nhằm ngăn chặn sự lan tỏa và nhân lên của virus. Trẻ suy giảm miễn dịch tế bào rất dãù bị nhiễm trùng virus lan tỏa như sởi, herpes simplex, varicella zoster, nhưng không bao giờ bị nổi ban đỏ đặc trưng ở da.
Bài viết cùng chuyên mục
Các đặc điểm chính của đáp ứng miễn dịch thu được
Tất cả đáp ứng miễn dịch bình thường, sẽ phai nhạt dần theo thời gian, để trả lại hệ miễn dịch ở trạng thái nghỉ ban đầu, tình trạng này gọi là hằng định nội môi.
Các con đường và cơ chế tái tuần hoàn và homing của tế bào lympho
Tế bào T nguyên vẹn có xu hướng homing, và tái tuần hoàn qua các cơ quan lymphô ngoại biên, để ở đó chúng sẽ nhận dạng, và đáp ứng với kháng nguyên.
Cytokin kích thích tạo máu
Các cytokin khác nhau kích thích, sự phát triển, và trưởng thành của nhiều dòng tế bào máu khác nhau.
Tính miễn dịch bẩm sinh và thu được
Các cơ chế của miễn dịch tự nhiên, cung cấp sức đề kháng ban đầu đối với nhiễm trùng, đáp ứng miễn dịch thu được đến muộn hơn, với sự hoạt hoá tế bào lymphô.
Một số hiệu quả thuận lợi của phản ứng tự miễn
Tìm được những chất tải, hiệu quả nhất cũng như các tá chất thích hợp, để có thể gây phản ứng ở mức độ mong muốn.
Khảo sát phức hợp miễn dịch
Hiện nay, trên thế giới, ta đã có bán những sản phẩm chuẩn, cho những phòng thí nghiệm miễn dịch đặc biệt chuyên khoa.
Khảo sát định tính immunoglobulin
Trong trường hợp không có bất thường chuỗi nặng, kháng huyết thanh chuỗi nhẹ tự do, tức không phản ứng với chuỗi nhẹ cố định vào chuỗi nặng.
Tế bào trình diện kháng nguyên
Tế bào hình sao được tìm thấy dưới lớp biểu bì, và trong đa số các cơ quan, ở đó chúng được đặt ở tư thế sẵn sàng để bắt giữ kháng nguyên.
Quá mẫn miễn dịch typ II
Phản ứng truyền máu đối với các thành phần khác của máu, như bạch cầu, tiểu cầu cũng có thể xảy ra, nhưng hậu quả của nó không nặng nề như phản ứng đối với hồng cầu.
Quá mẫn miễn dịch typ III
Kháng thể và kháng nguyên tạo thành phức hợp, phức hợp này hoạt hóa bổ thể đồng thời tác động gây giải phóng các amin hoạt mạch, làm tăng tính thấm thành mạch.
Tính tự miễn dịch
Sự hình thành một đáp ứng kháng thể bình thường, đối với đa số kháng nguyên protein đòi hỏi sự tham gia của 3 loại tế bào B, T, và tế bào trình diện kháng nguyên.
Đại cương thiếu hụt miễn dịch
Một đứa trẻ, hay một người lớn, sẽ bị nghi ngờ là thiếu hụt miễn dịch, khi trên cơ thể xuất hiện những nhiễm trùng lặp đi lặp lại, tồn tại kéo dài, trầm trọng hoặc bất thường.
Đại cương các tế bào chủ yếu của hệ thống miễn dịch
Tế bào tham gia vào đáp ứng miễn dịch thu được, bao gồm các lymphô bào đặc hiệu kháng nguyên, tế bào trình diện kháng nguyên.
Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Một phân tử IgM pentamer, kết hợp với kháng nguyên, là có thể cố định bổ thể, nhưng đối với IgG, thì phải có phân tử IgG được gắn với kháng nguyên ở vị trí gần nhau.
Sự hình thành phức hợp tấn công màng C5 9
Sau khi C5b gắn màng, C6 và C7 đến gắn vào C5b để tạo C5b67, C5b67 tác động với C8 để tạo C5b678, đơn vị này tạo phản ứng trùng hợp phân tử C9.
Thiếu hụt miễn dịch tiên phát
Đối với những trường hợp thiếu hụt kháng thể bẩm sinh, nhiễm trùng tái đi tái lại bắt đầu xuất hiện, trong khoảng thời gian từ tháng thứ 4 đến 2 tuổi.
Sự kết hợp kháng nguyên kháng thể
Khả năng trung hòa độc tố, và vi sinh vật của kháng thể, luôn phụ thuộc vào sự gắn kết chặt chẽ của chúng vào kháng thể.
Bệnh nguyên bệnh miễn dịch
Có những mô hình bệnh tự miễn ngẫu nhiên, trên động vật, rất có ích cho chúng ta nghiên cứu, đầu tiên là bệnh tuyến giáp tự miễn trên gà trống Obese.
Thiếu hụt miễn dịch thứ phát
Các thuốc ức chế miễn dịch, tác động lên rất nhiều khâu của chức năng tế bào, chức năng của lymphô bào và bạch cầu múi thường giảm.
Đại cương các kỹ thuật miễn dịch
Một số xét nghiệm sẽ trở nên vô ích, nếu chúng ta yêu cầu không đúng lúc, đúng chỗ, các phân chia sẽ giúp lâm sàng có được chỉ định thích hợp.
Các thành phần tế bào của hệ thống miễn dịch thu được
Có một nhóm tế bào lymphô thứ ba, là tế bào giết, đây là những tế bào tham gia vào hệ thống miễn dịch bẩm sinh, chống lại nhiễm trùng virus.
Khảo sát lymphô bào miễn dịch
Có hai loại phản ứng da in vivo, được dùng để phát hiện lympho bào T mẫn cảm đặc hiệu, đó là: thử nghiệm nội bì, dùng kháng nguyên tiêm vào lớp nội bì.
Cấu trúc phân tử của kháng thể
Các glycoprotein huyết tương, hoặc huyết thanh trước đây thường được phân chia theo tính chất hòa tan, của chúng thành albumin và globulin.
Miễn dịch chống vi khuẩn
Một số vi khuẩn xâm nhập qua đường niêm mạc, có thể tạo ra các protease để ly giải kháng thể IgA tiết, neisseria gonorrhea, neisseria meningitis.
Bằng chứng về tính chất gây bệnh của phản ứng tự miễn
Khi có được dòng động vật cảm thụ thích hợp rồi, ta còn có thể gây bệnh thụ động bằng cách truyền tế bào T mẫn cảm cho con vật khác.
